Cálcio muito além da função estrutural
O cátion cálcio (Ca++) na solução do solo segue o movimento do fluxo de massa na rizosfera promovido pela transpiração das plantas. Move-se pelo córtex dos pelos radiculares, sendo carregado no xilema com a transposição das membranas celulares por proteínas canais e transportadoras. Seguindo pelo fluxo transpiratório para os tecidos em crescimento, o Ca++ é distribuído pelo apoplasto (espaço da parede celular e lamela média), local de sua função estrutural, ou transpõe as membranas celulares para o simplasto (espaço contínuo – célula a célula – dos citoplasmas), local de sua função metabólica.
No apoplasto, além de celulose e hemicelulose, há proporções variáveis de pectinas, principalmente do ácido poligalacturônico na lamela média, cujos grupos carboxílicos (R – COO–) atuam como trocadores de cátions (Fig. 1). Isso confere ao apoplasto a capacidade de troca catiônia (CTC). Geralmente, a CTC das espécies dicotiledôneas é maior que a das espécies monocotiledôneas, e dos tecidos jovens é significativamente maior que a dos tecidos maduros, nos quais as cargas já foram saturadas pelo Ca++ (Wang et al., 2017; White et al., 2018).
Uma grande proporção do Ca++ total nos tecidos vegetais está ligada a CTC na lamela média, mas, a depender da disponibilidade, cátions trivalentes (Al+++) podem se ligar mais fortemente do que cátions divalentes (Ca++, Mg++), que por sua vez podem se ligar mais fortemente do que cátions monovalentes (K+) ou o próton H+ (Wu e Li, 2022).
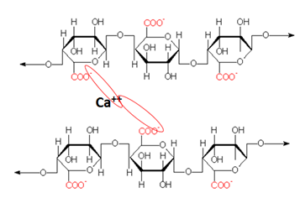
Figura 1. Representação do cálcio ligando as cargas negativas do ácido poligalacturônico na lamela média.
Portanto, a depender do suprimento de Ca++ nos tecidos em expansão, outros cátions ou o H+ podem ocupar essas cargas, alterando a conformação e integridade da lamela média. Assim, sintomas da deficiência de Ca++ como a queima dos bordos na alface e a podridão apical no tomate, estão relacionados à desagregação da lamela média nos tecidos jovens em expansão, com consequente desagregação também das membranas celulares, e o extravasamento e oxidação do conteúdo celular (Rengel, et al. 2023).
Além do Ca++ fixado no apoplasto, ocorre a transposição das membranas celulares pelo Ca++ para o simplasto. As células mantêm concentrações de Ca++ livre no citosol (matriz citoplasmática) de aproximadamente 50–200 nM (nanomolar), resultando em um gradiente químico (diferença de concentração) de Ca++ entre o citosol, o vacúolo que armazena a maioria do Ca++ no citoplasma, e o apoplasto. Com o reconhecimento de estímulos por proteínas nas membranas, as células modulam esses gradientes para gerar mudanças rápidas na concentração intracelular de Ca++, caracterizando sua função metabólica de sinalização (Behera et al., 2018).
Em sua função de sinalização, o Ca++ atua como mensageiro secundário da ação de hormônios vegetais e das respostas das plantas aos estímulos ambientais, como nos estresses bióticos e abióticos. Os sinais de Ca++ são resultado de uma complexa interação entre canais de influxo, efluxo e energia fornecida por ATP (adenosina trifosfato) (Ghosh, et al. 2022).
O padrão espaço-tempo do aumento da concentração de cálcio no citosol, e sua restauração aos níveis de repouso, ou seja, anteriores ao reconhecimento do estímulo, é chamado de assinatura de Ca++, que determina a especificidade da resposta correspondente ao estímulo que foi reconhecido (Wdowiak, et al. 2024). Esse processo pode ser amplificado para células adjacentes como “ondas de cálcio”, promovendo a difusão da sinalização (Lederer, 2025).
Assim, os aumentos transitórios na concentração de Ca++ no citosol promovem o aumento da concentração de Ca++ no núcleo com a passagem por poros e canais de Ca++ (Charpentier, 2018). O aumento da concentração no núcleo desencadeia a expressão gênica com a rápida regulação da transcrição, com o Ca++ ligando-se diretamente a fatores de transcrição (FTs), proteínas que controlam a transcrição do DNA em RNA mensageiro (mRNA), ou indiretamente por meio de “proteínas sensoras” ligantes de Ca++, como as calmodulinas (CaM) (Ghosh, et al. 2022; Ren et al., 2023) (Fig. 2), resultando em respostas de adaptação a estresses, com a síntese de enzimas, por exemplo.
O Ca++ e o CaM mediam as respostas das plantas induzidos por sinais de desenvolvimento e estresses ambientais, regulando a expressão gênica de diversas maneiras (Lederer, 2025). Portanto, a adaptação das plantas ao ambiente depende do estado nutricional de Ca++, muito além da sua função estrutural.
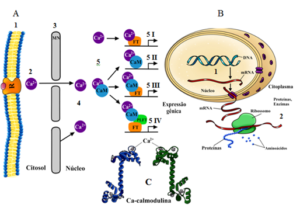
Figura 2. Representação esquemática da regulação da trasncrição mediados por Ca++ e Ca- calmodulina. A 1- Membrana plasmática e proteína receptora (R) ligada a uma molécula sinalizadora (ex. hormônios, eliciadores). 2 – Aumento da concentração de Ca++ no citosol (assinatura de Ca++ no citosol). 3 – membrana do núcleo (MN). 4 – Passagem do Ca++ pelos poros e canais na MN e aumento da concentração (assinatura de Ca++ no núcleo). 5 I – O Ca++ se liga diretamente ao fator de transcrição (FT) e modula sua atividade. 5 II – A calmodulina carregada com Ca++ (Ca++/CaM) se liga ao DNA e funciona como um FT. 5 III – O complexo Ca++/CaM interage com os FTs e modula sua função. 5 IV – O complexo Ca++/CaM regula indiretamente a transcrição, juntamente com FTs e proteínas de ligação ao fator de transcrição (PLFT), que funcionam como uma ponte entre o complexo Ca++/CaM e os FTs. B 1 – Transcrição, com a conversão de DNA em RNA mensageiro (mRNA). B 2- Síntese de proteínas codificada pelo mRNA organizando a sequência de aminoácidos. C – Complexo Ca++ calmodulina.
Fonte: adaptado de Kim et al., 2009; Ren et al., 2023; Lederer, 2025.
*Texto extraído do livro: “Interações entre fisiologia e nutrição de plantas – nutrientes e metabolismo”, a ser publicado pela Abisolo em 2026.
