Fatores climáticos relacionados ao enchimento de grãos
1 – INTRODUÇÃO
A performance da cultura de soja está fortemente associada e dependente das características ligadas ao crescimento e desenvolvimento. Vários aspectos morfológicos, como arquitetura da estrutura vegetativa, sistema radicular, formato e densidade de nervuras nas folhas, hábito de crescimento entre outros determinam o desempenho geral das plantas de soja. Por exemplo, a anatomia foliar e a arquitetura determinam a quantidade de luz interceptada, a capacidade fotossintética e a força fonte das plantas enquanto, a distribuição e densidade de nervura nas folhas direcionam o fluxo de fotoassimilados das regiões fonte para os drenos (Mathan; Bhattacharya; Ranjan, 2016).
Mas na prática, só esses fatores não são capazes de definir a produtividade dos cultivos de soja. De acordo com Buchanan; Gruissem; Jones (2000), as maiores perdas de produtividade de plantas cultivadas como é o caso da soja estão associadas a estresses abióticos. As reduções variam de 66 para o milho (Zea mays L.) até 81% para o sorgo (Sorghum bicolor). A explicação para esse processo está ligado a dificuldade de controlar eventos climáticos, que são instáveis e imprevisíveis.
Considerando todo o ciclo da cultura de soja, a fase reprodutiva é considerada a mais crítica, por ser extremamente sensível a estresses. Da mesma forma como ocorre na maioria das culturas, os estresses abióticos possuem maior peso na perda de produtividade.
Nesse período as plantas de soja possuem necessidade de síntese de grande quantidade de fotoassimilados, principalmente durante o enchimento de grãos. Nesta cultura a eficiência energética para a produção de grãos é baixa. São necessários 2 g de fotoassimilados para produzir 1 g de grão, enquanto que para outros cereais, apenas 1,3 a 1,4 g são utilizados (Sinclair; Wit, 1975). Esse fato é atribuído ao alto teor de lipídios e proteínas que são armazenados nos grãos que possuem elevado custo energético para serem produzidos, além de apresentarem elevada perda de energia pela respiração (Kakiuchi; Kobata, 2008).
Outro fator a ser considerado na fase reprodutiva da cultura de soja é a taxa de enchimento de grãos, cujo valor é variável de acordo a genética, disponibilidade de fotoassimilados, atividade do dreno e estádio fenológico. Normalmente, esses valores variam de 4,4 a 9,5 mg grão dia-1 do estádio R5 a R7 (Guffy et al., 1991). Dados atuais têm verificado valores médios de 8,3 mg grão dia-1, quando se utilizou a densidade de 20 plantas m-2 (El-Zeadani et al., 2014).
Abaixo são descritos alguns fatores climáticos de grande relevância no enchimento de grãos, principalmente quando estão em disponibilidades inadequadas.
2 – Deficiência hídrica
A deficiência hídrica na fase reprodutiva, especialmente no período de enchimento de grãos é considerado um dos fatores de maior impacto na produtividade de soja. Essa constatação já era verificada na década de 80, por Eck et al. (1987). De acordo com os autores, a baixa disponibilidade hídrica no período de enchimento de grãos ocasiona reduções de produtividade acima de 46%. Mais tarde, Brevedan; Egli (2003) relatam que plantas de soja submetidas a curtos períodos de deficiência hídrica (5 a 6 dias, com 40% da capacidade de campo) durante o enchimento de grãos podem reduzir a produtividade em até 39%.
Os principais componentes de produção afetados na cultuta de soja pela deficiência hídrica são: número de vagens, sementes por vagens, sementes por planta e massa de 1000 grãos (Ohashi et al., 2009; KobraeI et al., 2011), os quais perfazem os mais relevantes drenos de fotoassimilados (Nobuyasu et al., 2003). Brevedan e Egli (2003) relataram que curtos períodos de deficiência hídrica (em torno de 13 dias) durante o enchimento de grãos causam redução substancial na produtividade (39%), devido à redução no número de vagens e sementes. Esses autores também relataram diminuição na clorofila, fotossíntese e fluxo de carboidratos para os órgãos drenos.
A maior parte das perdas de produtividade está relacionada a alterações fisiológicas, como a redução na condutância estomática e fotossíntese, diminuição na expansão, celular e aumento na produção de espécies reativas de oxigênio, ERO (Jaleel et al., 2009). Em função disso ocorrem reduções do número de vagens, sementes por vagens, sementes por planta e massa de 1000 grãos (Ohashi et al., 2009; Kobraei et al., 2011), os quais perfazem os mais relevantes drenos de fotoassimilados (Nobuyasu et al., 2003). Além disso, o fluxo de carboidratos das raízes diminui causando a redução na absorção de nutrientes e do processo de fixação biológica de nitrogênio (Nunes, 2016).
O metabolismo de carboidratos e translocação de fotoassimilados para órgãos reprodutivos são os mais sensíveis a deficiência hídrica (Liu et al., 2004). Em função disso, aumentos nos teores de sacarose e hexose nas folhas são verificados, enquanto o amido diminui (Pelleschi et al., 1997). Isto evidência que nessas condições, o metabolismo celular prioriza a hidrólise de amido em “prol” da síntese de sacarose (Lemoine et al., 2013), para tentar manter a respiração foliar, além da osmorregulação. Outro efeito verificado se refere a falhas no sistema de condução de seiva no floema, devido ao aumento na viscosidade e alterações na permeabilidade da parede celular dos elementos crivados (Sevanto, 2014). Outro evento bastante comum em plantas com deficiência hídrica é a redução da atividade da enzima invertase (enzima que causa a quebra de sacarose). Essa enzima pode apresentar redução de até cinco vezes (Pinheiro et al., 2005). Tal comportamento reduz a capacidade de translocação de açúcares para órgãos drenos, que nesse momento é formado especialmente pelos grãos e vagens.
Atualmente, a maior parte de cultivares de soja lançadas no mercado possuem hábito de crescimento indeterminado. Essa característica pode ser relevante quanto a tolerância à deficiência hídrica, uma vez que as mesmas continuam crescendo após o início do florescimento mantendo o potencial de formação de novas flores e vagens após término do estresse (Villalobos-Rodrigues; Shibles, 1985). Tanaka e Shiraiwa (2009), buscando fortalecer esse conceito avaliaram as diferenças morfofisiológicas nas folhas de soja de ambos os hábitos de crescimento da soja. Os autores observaram que as plantas de hábito de crescimento indeterminado possuem maior densidade de estômatos e, consequentemente maior condutância estomática durante a deficiência hídrica, permitindo a planta tolerar mais intensamente esse estresse.
No entanto, as plantas podem utilizar mecanismos internos para minimizar os danos da deficiência hídrica. Dos quais, destaca-se o ajuste osmótico (aumento de prolina, glicina betaína e alguns açúcares solúveis) e a proteção de sistemas de membrana. Parece que o suprimento de alguns nutrientes também exerce um papel importante na tolerância das plantas de soja a esse estresse. Dentre eles estão os macronutrientes, potássio (K), cálcio, fósforo (P) e magnésio (Mg) os micronutrientes boro (B), Cobre (Cu) e zinco (Zn). Suas funções nas plantas são inúmeras, compreendendo a manutenção do balanço de cargas, transporte de elétrons, componentes estruturais, ativação de enzimas, ajuste osmótico e turgor celular (Waraich et al., 2011).
3 – Temperatura do ar
As plantas de soja submetidas a elevadas temperaturas causa diversos danos morfológicos e fisiológicos. Temperaturas acima de 30°C já ocasionam severas perdas de produtividade (Puteh et al., 2013), podendo ocasionar perdas de até 27%, quando esta atingir valores superiores a 35°C (Gibson; Mullen, 1996).
No metabolismo fisiológico, elevadas temperaturas atuam intensamente no metabolismo celular por afetar a cinética das reações químicas. Esses efeitos variam de acordo com o período diurno e noturno.
Para o processo fotossintético, baixas temperaturas reduzem a velocidade de transporte de elétrons e a fixação e redução do carbono. Esse processo aumenta rapidamente com o incremento da temperatura até atingir valores considerados ótimos. Para plantas C3, na sua grande maioria, esses valores situam-se entre 15 e 30°C (Larcher, 2004), sendo de 27°C para a cultura de soja. Outro efeito é a desidratação celular e desta forma a redução no fluxo de nutrientes e fotoassimilados no interior da planta
Outro efeito associado a elevada temperatura é a diminuição da capacidade de defesa da planta, devido à redução da atividade de enzimas antioxidantes, e o posterior acúmulo de ERO (Djanaguiraman; Prasad, Seppanen, 2010). Também é evidenciado o aumento na síntese de etileno e desintegração de estruturas celulares. Do mesmo modo, os danos morfológicos por elevadas temperaturas também são observados e relatados por Djanaguiraman et al. (2011). Os pesquisadores notaram que plantas de soja submetidas a elevadas temperaturas durante o florescimento apresentaram danos nos cloroplastos, dilatação na membrana dos tilacóides e a consequente redução na atividade fotossintética e senescência foliar. Esse mesmo efeito ocorre durante o enchimento de grãos e portanto, a sua influência na produção de fotoassimilados é similar ao que ocorre no florescimento.
Além disso, elevadas temperaturas afetam a performance de produtos aplicados via folha com objetivo de estímulo ou de proteção de plantas, pois causam desidratação de folhas e redução na translocação dessas moléculas.
4 – Radiação solar
A planta de soja por ser uma planta C3, possui baixa capacidade de utilização de luz (ponto de saturação luminosa de 1.800 µmol m-2 s-1 (Larcher, 2000), quando comparada com as demais plantas do grupo C4 [milho (Zea mays L) e cana de açúcar (Saccharum officinarum L.)]. Alterações de intensidades luminosas causam efeitos significativos em variáveis fisiológicas, afetando seu crescimento e desenvolvimento. Em condições de elevados níveis de radiação, a planta responde com alterações morfofisiológicas. Mesmo utilizando mecanismos de adaptação, as plantas de soja não apresentam uma proteção completa. E por isso, na maioria das vezes, o excesso de radiação solar ocasiona a produção de ERO, danos em clorofilas e incremento na fotorrespiração.
Em condições de baixos níveis de radiação solar, a cultura de soja apresenta vários mecanismos de adaptação. Em um vasto experimento Melges et al. (1989) buscou elucidar o efeito de níveis de luzia no crescimento e desenvolvimento da soja. O trabalho indicou que: (i) o número de folhas e vagens decresce em função do sombreamento; (ii) a taxa de acúmulo de fitomassa em cada órgão é tanto mais elevada quanto maior a densidade de fluxo de radiação solar, até a saturação luminosa; (iii) a taxa de acúmulo de fitomassa seca, devido ao sombreamento, foi menor nas raízes que nos outros órgãos; (iv) as plantas sombreadas apresentaram maior estatura, devido ao alongamento dos entrenós e aumento da dominância apical, (v) além disso, com o aumento do nível de sombreamento ocorreu maior retardamento na maturação.
A redução da disponibilidade de radiação solar verificado por Melges et al. (1989) em condições controladas é um estresse abiótico comum em algumas regiões do Brasil no período de estações chuvosas. Em um experimento recente em condições de campo, Ghassemi-Golezani, Bakhshi e Dalil (2015) confirmaram as constatações realizadas por Melges et al. (1989), especialmente, no que se refere a redução na dinâmica do fluxo de fotoassimilados em grãos, e o aumento no período efetivo de enchimento.
Em função disso, Fagan (2018, dados não publicados), desenvolveu um modelo que auxilia na explicação do efeito do sombreamento na fisiologia de plantas de soja (Figura 1). Para o pesquisador, durante a fase reprodutiva as plantas possuem a maior demanda de fotoassimilados para formação de órgãos reprodutivos. A redução da luminosidade, causa a limitação fotossintética diminuindo a produção de energia. Esse problema se agrava quando as plantas se encontram no período de enchimento de grãos (R5), quando a intensidade da atividade do dreno é máxima (os grãos em enchimento). Desta forma, com a baixa capacidade da fonte (folha) em suprir a alta atividade do dreno, ocorre o ajuste da relação fonte/dreno por meio do abortamento de vagens.
Normalmente, períodos nublados vêm acompanhados com chuvas em excesso e encharcamento de solo. O excesso de água no solo, além da capacidade de campo, diminui os níveis de oxigênio no solo, assim, ocorre acúmulo de ACC (ácido 1-carboxílico-1-aminociclopropano) nas raízes das plantas que são transportados via xilema para a parte aérea. Na parte aérea, com a disponibilidade de oxigênio, o ACC é convertido a etileno pela atividade da ACC-oxidase, (Fagan et al., 2015), ocasionando senescência de folhas, flores e vagens.
Também é importante salientar que, em plantas vasculares, o boro e o cálcio movem-se via corrente transpiratória. Em dias nublados, devido à redução no fluxo transpiratório, a disponibilização de Ca e B nas folhas, flores e vagens do estrato superior diminui. Como esses nutrientes são imóveis no floema, não ocorre a remobilização dos mesmos das regiões mais velhas para esses órgãos. Esse processo acentua ainda mais a abscisão de folhas, flores e vagens.
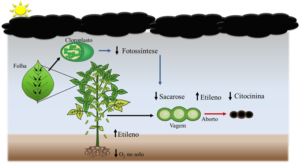
Figura 3.4. Possível modelo fisiológico envolvido no processo de aborto de vagens de soja submetidas a baixa luminosidade (elevada nebulosidade) e excesso de água no solo. Fagan (2018, dados não publicados).
No enchimento de grão, a redução da luminosidade no terço inferior é um dos principais fatores associados a senescência das folhas baixeiras e consequentemente, a redução no enchimento de grãos. Mas porque a perda destas folhas tem tanta influência no enchimento de grãos? Primeiramente precisamos entender que em condições de diminuição de luminosidade, as folhas apresentam menor taxa do fluxo transpiratório resultando na redução do fluxo de citocininas (Pons; Jordi, 1998). Associado a isso, a baixa intensidade de luz causa uma redução na fotossíntese bruta até o momento em esta torna-se inferior a respiração foliar. Nesse momento, inicia-se a expressão de genes que codificam proteínas fotossintéticas e enzimas relacionadas a formação de clorofila (Thomas, 1978). Esse processo inicia pelo fitocromo, que atua como receptor de luz vermelha e vermelha extrema. A teoria mais aceita afirma que, quando a intensidade da luz cai abaixo do ponto de compensação de luz, ocorre o balanço negativo de carbono que, por sua vez, desencadeia a senescência (Boonman et al., 2006).
5 – Conclusões
É importante destacar que a produtividade da cultura da soja não é definida apenas pelo período de enchimento de grãos, mas que depende de um planejamento adequado quanto ao manejo de solo, proteção de plantas e nutrição. Contudo pode-se perceber através dessa breve resenha, que o enchimento de grãos é extremamente sensível a estresses climáticos, uma vez que nessa fase a necessidade de energia é elevada e que precisa ser distribuídas para os grão em um período relativamente curto (20 a 30 dias).
Evandro Binotto Fagan
2020 - Abisolo
