Brassinosteroides: moduladores fisiológicos de plantas sob estresses abióticos
O ambiente de cultivo tem um impacto significativo na produtividade das plantas cultivadas. Diversos estresses, como deficiência hídrica, temperatura extrema, salinidade, luminosidade, pragas e doenças, entre outros, afetam as plantas de diferentes maneiras. Para que as plantas consigam sobreviver, é necessário levar em consideração algumas características, como a severidade, duração, número de exposições e combinação de vários estresses. Além disso, o efeito desses estresses varia de acordo com o órgão do vegetal que foi exposto, o estádio de desenvolvimento da planta e o genótipo. Se a planta conseguir resistir, ocorre a sobrevivência e o crescimento, no entanto, quando a suscetibilidade é inevitável, a senescência ocorre (Buchanan; Gruissem; Jones, 2015, Figura 1).
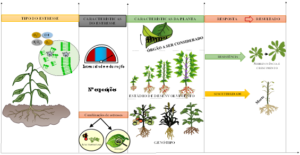
Figura 1. Fatores que influenciam a resposta das plantas ao estresse. i) Tipo de estresse; ii) característica do estresse (Intensidade, duração, número de exposições ao estresse e combinações de estresses); iii) características da planta (órgão que está sendo afetado, estádio de desenvolvimento da planta e genótipo), iv) resposta e resultado que considera a resistência que leva a sobrevivência e crescimento e a suscetibilidade que ocasiona a senescência da planta.
Fonte: Adaptado de Buchanan; Gruissem; Jones (2015).
As alterações no metabolismo e no desenvolvimento das plantas induzidas por estresses ambientais estão intimamente associadas a mudanças coordenadas nos padrões de expressão gênica. Essas respostas são desencadeadas quando o estresse é inicialmente percebido em nível celular, funcionando como um sistema de “sensores internos” capazes de detectar desvios na homeostase. Uma vez reconhecido, o sinal é amplificado por cascatas de transdução que propagam a informação entre células e tecidos, integrando respostas locais em ajustes sistêmicos que modulam o crescimento, o desenvolvimento e a capacidade reprodutiva da planta. A intensidade, a duração e a frequência do estresse atuam como reguladores do “ritmo” da resposta, determinando sua magnitude e velocidade.
Entre os múltiplos ajustes metabólicos ativados sob condições adversas, as alterações no metabolismo hormonal destacam-se como um dos mecanismos mais sensíveis e decisivos. Nesse contexto, a via de sinalização do ácido abscísico (ABA) é amplamente reconhecida como uma rota central na percepção e na resposta aos estresses ambientais, especialmente aqueles relacionados à seca, salinidade e temperatura e radiação extremas. No entanto, evidências consistentes demonstram que um hormônio ainda pouco conhecido denominado de brassinosteroides (BRs) também exercem papel estratégico nesse processo, atuando tanto por meio de interações diretas (“crosstalk”) com a sinalização do ABA quanto de forma independente. Assim, os BRs funcionam como um “regulador de intensidade fisiológica”, permitindo que a planta ajuste seu metabolismo de maneira equilibrada, evitando tanto a paralisação do crescimento quanto o gasto energético excessivo sob condições adversas (Zhu et al., 2016).
Embora o interesse na utilização de BRs seja relativamente recente, sua descoberta remonta a mais de quatro décadas (Peres et al., 2019). Esse hormônio pertence à classe dos esteroides polihidroxilados e regulam uma ampla gama de processos fisiológicos, incluindo divisão, alongamento e ciclo celular, morfogênese, reprodução, senescência e respostas ao estresse (Oh et al., 1998). Inicialmente, os BRs foram identificados em grãos de Brassica napus devido à sua capacidade de promover alongamento e divisão celular, o que levou à proposição de uma nova classe de hormônios vegetais, denominada brassinas (Mitchell et al., 1970). Posteriormente, o brassinolídeo (BL), considerado o BR biologicamente mais ativo, foi isolado em 1979 a partir de grãos de pólen de canola, em um processo que ilustra a extrema eficiência fisiológica desses compostos mesmo em concentrações traço (Grove et al., 1979).
Diversos mecanismos explicam como a sinalização mediada por BRs contribui para a adaptação das plantas ao estresse. Entre os principais destacam-se: (i) o ajuste fino da maquinaria transcricional, permitindo respostas gênicas proporcionais à intensidade do estímulo estressor (Ye et al., 2017); (ii) a ativação de sistemas antioxidantes (sistema enzimática e não enzimático), que funcionam como um “escudo molecular” contra o excesso de espécies reativas de oxigênio (ERO) geradas em condições adversas (Zou et al., 2018) além da síntese de proteínas de choque térmico; e (iii) a estimulação da produção de osmoprotetores (prolina), compostos que preservam a integridade celular e o equilíbrio osmótico, especialmente sob déficit hídrico e salinidade (Fàbregas et al., 2018).
A interação entre BRs e ABA exerce influência direta sobre a expressão de genes relacionados ao estresse, ao desenvolvimento vegetal e à germinação de sementes de forma independentes (Shah et al., 2019; Taiz et al., 2024). Evidências experimentais indicam que o tratamento com BRs pode atenuar limitações fisiológicas na germinação, inclusive em mutantes com deficiência na biossíntese de giberelinas (Steber e Mccourt, 2001), além de reduzir os efeitos inibitórios do ABA em Arabidopsis (Divi e Krishna, 2010). Resultados semelhantes foram observados em Brassica napus sob estresse salino e frio, por estimular a síntese de proteínas de choque térmico (HSP) (Kagale et al., 2006). Um dos mecanismos centrais do BRs na tolerância ao frio envolve o acúmulo das formas ativas não fosforiladas dos fatores de transcrição BZR1 e BES1, que promovem a indução de genes da família CBF (“C-repeat/Dehydration-Responsive Element Binding Factors”), especialmente CBF1 e CBF2, reconhecidos como reguladores positivos das respostas ao frio. Essa ativação fortalece a expressão de genes responsivos ao congelamento, contribuindo para o aumento da tolerância celular a temperaturas baixas (Li et al., 2017).
Em algodão, Chakma e colaboradores observaram que a aplicação de 2 µM de 24-epibrassinolídeo (EBR) aumenta a germinação de sementes sob temperaturas elevadas (70 a 75 °C), além de mitigar a inibição da germinação induzida por ABA. A capacidade dos brassinosteroides de estimular o crescimento de plântulas sob diferentes condições de estresse indica que o priming de sementes de algodão com BR pode favorecer um estabelecimento inicial mais rápido e consistente. O mesmo tratamento favoreceu a abertura dos cotilédones e o desenvolvimento de raízes laterais, sugerindo que a pré-embebição de sementes com EBR pode atuar como um “impulso inicial” para o estabelecimento precoce das plântulas em ambientes estressantes (Chakma et al., 2021).
Os BRs também participam da regulação do crescimento sob baixa luminosidade, atuando em conjunto com criptocromos e auxinas no alongamento do hipocótilo e na evitação de ambientes sombreados (Taiz et al., 2024). Além disso, esses hormônios estão presentes em elevadas concentrações nos grãos de pólen, onde promovem a germinação e o crescimento do tubo polínico em Arabidopsis thaliana, para o sucesso reprodutivo (Vogler et al., 2014). No gametófito feminino, os BRs são igualmente essenciais para a iniciação das divisões mitóticas do megásporo, sendo indispensáveis ao desenvolvimento do megagametófito (Taiz et al., 2024).
Do ponto de vista aplicado, o uso de BRs tem se mostrado promissor para o aumento da produtividade vegetal sob estresses abióticos (Özdemir et al., 2004; Upreti e Murti, 2004; Ahammed et al., 2020). Um modelo fisiológico proposto por Ahammed e colaboradores descreve que os BRs induzem a expressão do gene RBOH1 (“Respiratory Burst Oxidase Homolog 1”) responsável pela produção controlada de ERO no apoplasto. Esses sinais redox atuam como mensageiros secundários que ativam fatores de transcrição e genes associados à síntese de proteínas protetoras, integrando-se à sinalização mediada por MAPKs em um mecanismo de retroalimentação positiva (Ahammed et al., 2020).
Em soja, Zhang e colaboradores demonstraram que a aplicação foliar de brassinolídeo no florescimento atenua os efeitos da deficiência hídrica, promovendo aumentos na eficiência quântica do fotossistema II, na atividade da enzima RuBisCO, no conteúdo relativo de água, nos teores de açúcares solúveis e prolina, além de intensificar a atividade das enzimas antioxidantes POD e SOD (Zhang et al., 2008). Em experimento similar, Yadava e colaboradores observaram que a aplicação de 24-epibrassinolídeo (1 µM) em plantas de milho submetidas a elevadas temperaturas (em torno de 48 °C) aumentou a atividade das enzimas CAT, POD e SOD e os teores de proteína total que proporcionou maior proteção de membranas celulares aos danos oxidativos (Yadava et al., 2016). No metabolismo fotossintético, níveis elevados de BRs estão associados ao aumento das taxas de carboxilação da Rubisco e do transporte de elétrons nos fotossistemas, além da mitigação da fotoinibição por meio do fortalecimento do sistema antioxidante (Ahammed et al., 2015; Shu et al., 2016). Adicionalmente, os BRs modulam o fluxo eletrônico em cloroplastos e mitocôndrias via ativação de oxidases alternativas, dissipando o excesso de energia e reduzindo a formação de ERO (Deng et al., 2007).
Os BRs também influenciam o metabolismo do nitrogênio. Em tomateiro, a aplicação de 24-epibrassinolídeo aumenta a atividade de enzimas-chave do metabolismo nitrogenado e de bombas de íons, contribuindo para a manutenção do gradiente eletroquímico sob estresse (Shu et al., 2016). Em milho, Xing e colaboradores demonstraram que a aplicação exógena de EBL regula a expressão de transportadores de nitrato (NRT), promovendo maior crescimento radicular, acúmulo de biomassa e teor de nitrogênio nos tecidos (Xing et al., 2022). Em função disso, plantas submetidas a baixos teores de N no solo, podem usar mais eficientemente esse nutriente na presença dos BRs.
Por fim, os BRs regulam o crescimento radicular de maneira dependente da concentração, controlando o tamanho das células meristemáticas e estimulando o transporte polar de auxina. Ao aumentar a expressão de proteínas PIN, os BRs favorecem a formação de raízes laterais (Figura 2), atuando em sinergia com açúcares como a glicose, que funcionam como sinais metabólicos integradores do desenvolvimento radicular.

Figura 2. Efeito da aplicação de brassinolídeos em sementes de soja comparado a outros enraizadores consolidados no meracado (EA, Ascophyllum nodosum L.).
Em síntese, os BRs atuam como hormônios centrais na integração das respostas ao estresse, conectando sinais ambientais às respostas fisiológicas, metabólicas e estruturais da planta (Figura 3). Sob condições de estresse abiótico, os BRs desencadeiam uma resposta altamente coordenada, envolvendo múltiplos processos celulares e diferentes órgãos vegetais. Nesse contexto, os BRs: (1) ativam o sistema antioxidante, estimulando enzimas como a CAT, APX e SOD, o que reduz o acúmulo de espécies reativas de oxigênio; (2) promovem o crescimento e a plasticidade do sistema radicular, favorecendo a absorção de água e nutrientes, especialmente nitrogênio [transportadores de nitrato e enzimas de assimilação, (nitrato e nitrito redutase)] em ambientes adversos; (3) modulam as trocas gasosas, ajustando a assimilação de CO₂ e a perda de água, com impacto direto sobre a eficiência fotossintética por atuar em conjunto com o ABA; (4) contribuem para a atenuação da toxicidade por metais pesados, preservando a homeostase metabólica; (5) interagem funcionalmente com o ABA, regulando a abertura estomática e o balanço hídrico foliar; (6) aumentam a termotolerância (aumenta expressão de proteínas de choque térmico, HSP), promovendo a estabilização de membranas, proteínas e complexos fotossintéticos; e (7) atuam no alongamento do hipocótilo em dicotiledôneas. De forma integrada, os BRs funcionam como um verdadeiro “orquestrador fisiológico”, coordenando a homeostase redox, o equilíbrio hídrico, a eficiência fotossintética, o ajuste osmótico e a resiliência metabólica. Essa integração permite que a planta mantenha o crescimento, o funcionamento fisiológico e a produtividade, mesmo sob condições ambientais limitantes.
Parte inferior do formulário
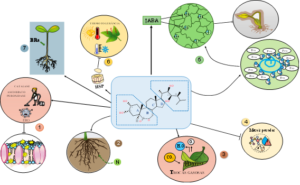
Figura 3. Brassinosteroídes (BRs) e respostas de plantas ao estresse. A sinalização por BRs ativa sistemas antioxidantes (CAT, SOD e APX), preserva a integridade de membranas, modula a abertura estomática em interação com ABA, mantém a eficiência fotossintética e estimula o crescimento radicular e o acúmulo de osmólitos (ex.: prolina), conferindo maior resiliência fisiológica sob estresses abióticos.
Evandro Binotto Fagan
Marina Rodrigues dos Reis
2026 - Abisolo
