A importância da nutrição mineral para a resistência das plantas às doenças
As culturas agrícolas estão expostas aos mais diferentes tipos de estresses, sejam esses de origem abiótica e ou biótica, durante praticamente todo o período de cultivo. Dentre os estresses de natureza biótica, a ocorrência de doenças, as quais são causadas por patógenos (ex. bactérias, fungos, nematoides, oomicetos e vírus) de diferentes estilos de vida (biotróficos – dependentes do tecido foliar para nutrição e, portanto, não causam necrose; necrotróficos – causadores de clorose seguida de necrose devido a ação de toxinas e enzimas hidrolíticas e os hemibiotróficos – inicialmente biotróficos, mas rapidamente comportam-se como necrotróficos), ocupa papel de destaque. As doenças contribuem significativamente para reduzir tanto a quantidade quanto a qualidade da produção. Muitas pessoas indagam por que ocorre aumento no preço do tomate, um dos itens que faz parte das refeições da maior parte dos brasileiros, em determinadas épocas do ano fazendo com que o índice nacional de preços ao consumidor amplo acelere, acarretando aumento da inflação. O aumento na intensidade das doenças devido às condições de maior umidade relativa e temperaturas mais amenas em decorrência das chuvas é uma das explicações mais plausíveis.
Os patógenos possuem diferentes estratégias para infectar os seus hospedeiros e obter os nutrientes necessários para garantir a reprodução e, assim, perpetuar a sua espécie na natureza. Várias alterações fisiológicas e bioquímicas ocorrem nas plantas infectadas pelos patógenos. A fotossíntese é o principal processo fisiológico afetado pelos patógenos. Os patógenos manipulam o metabolismo dos carboidratos nas plantas infectadas afetando tanto o crescimento quanto a produção. A absorção e a translocação de água e nutrientes nas plantas infectadas por patógenos causadores de murcha vascular e podridão das raízes sofrem drástica mudança. O estresse oxidativo nos tecidos dos hospedeiros infectados culmina com a produção excessiva de espécies reativas de oxigênio que maximizam o dano celular. A infecção por patógenos também ocasiona alterações no balanço hormonal da planta, na translocação dos fotoassimilados e nas atividades das enzimas associadas com o metabolismo do carbono.
Resistência é a capacidade das plantas em evitar ou atrasar a penetração e ou a subsequente colonização dos seus tecidos pelo patógeno utilizando-se das mais diversas estratégias de defesa. Todas as plantas apresentam um nível de resistência a qual denominamos de basal. Os mecanismos de defesa, estruturais e ou de natureza bioquímica, denominados de constitutivos (ex. camada de cera e de cutícula mais espessas, anatomia das células guardas dos estômatos, abundante presença de tricomas, estrutura e organização dos vasos condutores e presença de compostos de natureza antimicrobiana) ou pós-formados em resposta à infecção (ex. papilas, lignificação da parede celular, formação de camada de cortiça, zonas de abscisão, formação de tiloses e aumento na concentração de compostos fenólicos e fitoalexinas) garantem a defesa das plantas contra os seus patógenos. Nesse contexto, vários componentes de resistência da planta devem ser avaliados para confirmar a redução nos sintomas da doença: o tempo para o aparecimento dos sintomas da doença (período de incubação) e para que o patógeno se reproduza no tecido vegetal (período latente) são prolongados, as lesões expandem-se mais lentamente durante o desenvolvimento da doença e são de menor tamanho e o número de esporos produzidos nessas lesões é drasticamente reduzido. Para que as plantas possam expressar o seu máximo nível de resistência basal, faz-se necessário um maior fluxo de carbono via rotas do ácido chiquímico e dos fenilpropanóides para que os compostos de defesa sejam formados. Por exemplo, o manganês (Mn) é de extrema importância para que a produção de compostos fenólicos via rota dos fenilpropanóides ocorra com a melhor eficiência possível. Mecanismos de defesa pós-formados devem ser efetivos, no tempo e espaço, para conter a infecção pelo patógeno. Assim, a intensidade da doença será reduzida em função do nível de expressão da resistência. Em outras palavras, quanto maior o número de estratégias de defesa que estão ocorrendo no sítio de infecção do patógeno maior será a resistência da planta e, consequentemente, menor a intensidade da doença. Com a participação efetiva dos diferentes mecanismos de defesa na tentativa de reduzir a colonização dos patógenos nos tecidos dos seus hospedeiros é que iremos controlar as doenças com menor dependência da aplicação de fungicidas, bactericidas, nematicidas e até mesmo de inseticidas ao considerarmos que alguns patógenos são transmitidos por insetos-praga. O estado fisiológico das plantas é crucial para que a resistência basal em resposta a infecção por um determinado patógeno seja expressa da maneira mais robusta possível. Assim, fatores que afetam o metabolismo primário da planta tais como baixa assimilação de carbono, baixa luminosidade, desequilíbrio nutricional, déficit hídrico e problemas no desenvolvimento do sistema radicular passam a impactar a resistência basal.
Ao falarmos das estratégias de defesa que as plantas possuem para conter a infecção por um determinado patógeno não devemos nos esquecer da importância de se manter o correto estado nutricional delas. A nutrição mineral é a prática cultural de maior aplicabilidade pelos agricultores para o controle das doenças que acometem as culturas de grande importância econômica. A resistência das plantas em resposta à infecção por patógenos pode ser potencializada pela concentração de um determinado nutriente no tecido vegetal, bem como pela interação entre os nutrientes. Considerando a dificuldade de se prever a ocorrência de uma determinada doença e o seu progresso durante o ciclo da cultura pela dependência do fator ambiental, é mais desafiador ainda manter as plantas num estado nutricional que seja desfavorável aos patógenos durante os seus processos infecciosos. Cada nutriente colabora, direta ou indiretamente, para que a resposta das plantas à infecção pelos patógenos através da ativação de rotas bioquímicas de defesas seja a mais eficiente possível. Para cada interação planta-patógeno, os nutrientes estão envolvidos no funcionamento das diferentes rotas metabólicas que culminam com a produção de compostos de natureza antimicrobiana, lignina e ativação dos genes que codificam para enzimas de defesa (ex. fenilalanina amônia-liase (FAL), quitinase (QUI), β-1,3-glucanase (GLU), polifenoloxidase (PFO), peroxidase (POX) e lipoxigenase (LOX)) e do metabolismo antioxidativo que irão afetar o processo infeccioso de cada patógeno. Não devemos desconsiderar o efeito direto dos nutrientes no metabolismo dos patógenos, principalmente os que habitam o solo e infectam as plantas pelo sistema radicular devido, principalmente, a alteração do pH da rizosfera. Devemos estar conscientes de que a nutrição mineral deve ser vislumbrada holisticamente considerando que a absorção de um determinado nutriente é muitas das vezes influenciada por outro nutriente. Além disso, cada nutriente afeta a fisiologia das plantas de diferentes maneiras e, assim, propicia condições que pode favorecer ou não a infecção pelos patógenos.
Considerando a importância da nutrição mineral como uma alternativa para o controle das doenças das plantas no contexto de uma Agricultura sustentável, umas das linhas de pesquisa do Laboratório da Interação Planta-Patógeno (LIPP) do Departamento de Fitopatologia na Universidade Federal de Viçosa busca elucidar como os nutrientes, incluindo o silício, atuam na resistência das plantas quando infectadas por patógenos utilizando-se de análises fisiológicas, bioquímicas e moleculares.
Alguns estudos conduzidos no LIPP investigaram a hipótese de que plantas de arroz e trigo supridas com altas doses de magnésio (Mg) apresentariam redução nos sintomas de doenças foliares e teriam a fotossíntese preservada culminando, assim, em mais energia para ser alocada para as rotas de defesa das plantas. Para a escaldadura do arroz (Monographella albescens), o aumento na concentração foliar de Mg para as plantas supridas com 1,5 mM desse nutriente em comparação com a dose de 0,5 mM resultou em menores lesões da escaldadura em associação com maior concentração de compostos fenólicos e maiores atividades das enzimas FAL, PFO, POX e LOX. Além disso, um maior suprimento de Mg garantiu a preservação da capacidade fotossintética nas folhas infectadas por M. albescens, as quais apresentaram reduzida disfunção bioquímica a nível dos cloroplastos conjuntamente com uma maior concentração de pigmentos fotossintéticos. Já para a brusone (Pyricularia oryzae) do trigo, a severidade da doença aumentou de 11 para 40% para as plantas crescidas em solução nutritiva com as concentrações de 0,25 e 4 mM Mg, respectivamente. Na dose de 4 mM Mg, as plantas infectadas apresentaram redução na fotossíntese, condutância estomática, transpiração e concentração de pigmentos fotossintéticos. De um outro lado, plantas de arroz crescidas em solução nutritiva com diferentes concentrações de Mg (0,25; 2,5 e 4 mM) tornaram-se mais resistentes à mancha parda (Bipolaris oryzae) na maior dose desse nutriente devido as maiores atividades das enzimas FAL, PFO e POX. Fisiologicamente, as plantas crescidas na dose de 4 mM Mg apresentaram melhor condutância estomática e, consequentemente, maior disponibilidade de CO2 nos sítios de carboxilação, além de não sofrerem perdas significativas na concentração de pigmentos fotossintéticos (clorofila a, clorofila b e carotenoides) durante o processo infeccioso de B. oryzae. Uma maior concentração de Mg nas bainhas de plantas de arroz crescidas em solução nutritiva contendo 0,50 mm Mg reduziu significativamente a expansão das lesões da queima-das-bainhas (Rhizoctonia solani).
No LIPP também foi investigado o efeito do cálcio (Ca) na potencialização da resistência do trigo à brusone (P. oryzae). As plantas de trigo foram cultivadas em solução nutritiva contendo três concentrações de Ca (0,26, 2,6 e 5 mM) e não inoculadas ou inoculadas com P. oryzae. A concentração foliar de Ca aumentou de 1,3 para 4% e a severidade da brusone reduziu de 37 para 8% da menor para a maior concentração de Ca, respectivamente. O nível de expressão dos genes marcadores para as rotas de defesa mediadas pelos hormônios ácido salicílico (AS) e ácido jasmônico (AJ) (PR-1 e aleno óxido sintase, respectivamente), bem como para FAL, GLU e QUI, foi maior para as plantas infectadas e supridas com 5 mM Ca em comparação com a dose de 0,26 mM Ca. Somado ao conhecido efeito do Ca no fortalecimento da parede celular e na inibição de enzimas secretadas por patógenos fúngicos, os resultados desse estudo indicaram o potencial do Ca em potencializar a resistência do trigo à infecção por P. oryzae.
Para a mancha parda do arroz, a severidade da doença aumentou em função da maior disponibilidade de zinco (Zn) às plantas. Para a mela (R. solani) da soja, não houve relação significativa entre as maiores concentrações foliares de Zn e Mn com o desenvolvimento da doença.
Altas concentrações foliares de nitrogênio e potássio (K) resultaram em menor severidade da mancha parda em arroz. Maior concentração de K nas bainhas das plantas de arroz cultivadas em solução nutritiva contendo 100 mM K resultaram em menores lesões da queima-das-bainhas. No caule de plantas de manga supridas com solução de 0,5 M de cloreto de potássio, a extensão da área necrótica causada pelo fungo Ceratocystis fimbriata foi menor devido a maior concentração de K.
A ferrugem asiática da soja (Phakopsora pachyrhizi) tem causado perdas significativas na produção de soja e a aplicação de níquel (Ni) pode contribuir para aumentar a resistência das plantas em resposta à infecção pelo fungo. Em um estudo conduzido no LIPP, a aplicação foliar de Ni reduziu a severidade da doença em 38% em comparação com as plantas que não receberam esse micronutriente (Fig. 1). A redução nos sintomas foi associada com maior estímulo do sistema de defesa das plantas: houve maior concentração de lignina nos folíolos, além de maior atividade da GLU e expressão dos genes de defesa CHI, FAL e PR-1.
Vários estudos realizados no LIPP relataram o potencial do silício (Si) como estratégia de controle de várias doenças de plantas, especialmente ocorrendo em monocotiledôneas (ex., arroz, aveia, cevada e trigo). Nos inúmeros patossistemas investigados, a deposição e a polimerização do Si abaixo da cutícula e parede celular das células do tecido foliar evitou a penetração dos patógenos e ou atrasou o ingresso deles explicando, pelo menos em parte, a redução na severidade das doenças estudadas. Paralelamente, um atraso na penetração dos patógenos e até mesmo redução na colonização dos tecidos dos seus hospedeiros permitiu com que as plantas tivessem mais tempo para ativar, de forma mais eficiente, os seus mecanismos de defesa tais como aumentar a produção de compostos fenólicos, fitoalexinas e lignina e maximizar a atividade das enzimas de defesa FAL, GLU, QUI, PFO e PFO. Acredita-se que a deposição do Si na parede celular das células do tecido foliar afetaria a chegada de efetores produzidos pelos patógenos nos sítios de ação fazendo com que os mecanismos de defesa das plantas se tornassem mais eficientes a contar também que no interior das células, o Si solúvel estaria potencializando esses mecanismos de defesa. A deposição de material de natureza fenólica ao redor das hifas do fungo Pyricularia grisea colonizando as células do mesofilo em folhas de plantas de arroz supridas com Si afetou o crescimento delas (Fig. 2) explicando, assim, a redução na expressão dos sintomas da brusone em comparação com as plantas não supridas com esse elemento (Fig. 3).
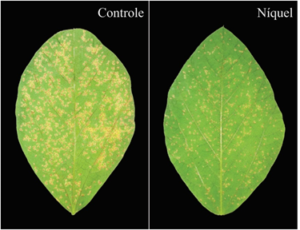
Figura 1. Sintomas da ferrugem Asiática da soja, causada pelo fungo Phakopsora pachyrhizi, nos folíolos obtidos de plantas de soja pulverizadas com água (controle) ou com níquel.
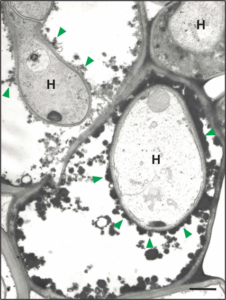
Figura 2. Hifas do fungo Pyricularia grisea, causador da brusone, infectando células do mesofilo foliar de plantas de arroz supridas com silício. Nota-se a deposição de material eletrodenso e osmiofílico (cabeça de setas na cor verde), indicativo da presença de compostos fenólicos, ao redor das hifas. Esses compostos fenólicos afetam o crescimento das hifas por exercerem uma natureza antimicrobiana evitando a colonização das células adjacentes. H = hifas do fungo. Barra = 1 µm.
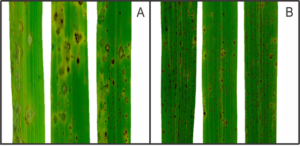
Figura 3. Sintomas da brusone, causada pelo fungo Pyricularia grisea, nas folhas de plantas arroz não supridas (A) ou supridas com silício (B).
Fabrício Ávila Rodrigues
2020 - Abisolo
