Poliaminas: moléculas bioativas derivadas de L-aminoácidos
Os L-aminoácidos formam proteínas e são precursores de moléculas com funções biológicas essenciais para as plantas, como enzimas, hormônios, polímeros da parede celular, participam do transporte de nutrientes e da adaptação das plantas aos estresses abióticos (Mógor & Mógor, 2022).
Entre as inúmeras moléculas bioativas derivadas de L-aminoácidos, as poliaminas (PAs), cuja biossíntese depende dos aminoácidos L-arginina, L-ornitina e L-metionina, estão presentes em todas as células vegetais, participando das transições dos estádios fenológicos, do crescimento e da senescência, com sua distribuição e concentração mudando de acordo com o desenvolvimento e adaptação das plantas ao ambiente (Pál et al., 2021).
As principais formas livres de PAs são a putrescina (Put), sendo catalisada a partir da L-arginina e L-ornitina; a espermidina (Spd) catalisada a partir da Put, e a espermina (Spm) catalisada a partir da Spd, com a participação da molécula reguladora S-adenosilmetionina derivada do aminoácido L-metionina (Chen et al., 2019).
Por serem hidrossolúveis e de baixo peso molecular, as PAs se difundem pelo tecido vegetal desempenhando múltiplas funções metabólicas, como divisão e alongamento celulares, embriogênese, diferenciação floral, desenvolvimento do cloroplasto, senescência foliar (Hasan et al, 2021; Pál et al., 2021), participam no transporte de cátions e homeostase (equilíbrio) celular de Ca++, K+ e Mg++ (Pottosin et al., 2021; Pathak & Dudhagi, 2021) e nas respostas aos estresses abióticos, estimulando a capacidade antioxidante das plantas (Seo et al., 2019).
Entre as inúmeras funções metabólicas das PAs, de maneira geral Spd e Spm estão relacionadas à promoção do crescimento das plantas, enquanto a Put está associada aos processos de maturação e senescência (Xie et al., 2014; Anwar et al., 2015; Ahmed et al., 2017).
Trabalhos recentes relacionam a bioatividade de fontes de L-aminoácidos aplicados às plantas com o metabolismo das PAs, a exemplo da aplicação foliar de um hidrolisado proteico obtido por processo enzimático em plantas de alface, que alterou o metabolismo das PAs, promovendo o aumento no teor de Spm nas folhas, estimulando o crescimento das plantas (Mógor et al., 2018).
O processo de brotação das gemas dos colmos de cana-de açúcar também foi relacionado ao metabolismo das PAs, com a imersão de mini-toletes em solução contendo biomassa rica em L-aminoácidos apresentando aumento significativo na brotação e crescimento dos brotos, sendo esses efeitos relacionados ao aumento da Spd e redução do teor de Put nos brotos, alterando dessa forma o perfil das PAs para estimular as brotações (Mógor et al., 2022). Incrementos nos níveis de Spd e a redução de Put podem melhorar o transporte da sacarose (Luo et al., 2019), justificando a maior redistribuição de açúcares não redutores dos mini-toletes para as gemas durante a brotação, em função da imersão na solução com a biomassa rica em L-aminoácidos (Mógor et al., 2022).
Além da promoção do crescimento vegetal, alternativas para mitigar os efeitos dos estresses abióticos são prioritários como temas de pesquisa, entre os quais se destacam os recentes estudos dos efeitos das PAs na adaptação das plantas ao ambiente.
Para superar o efeito dos estresses ambientais, o acúmulo de diferentes osmólitos (carboidratos, glicina-betaína, prolina e outros aminoácidos) e a atividade de enzimas antioxidantes são fatores adaptativos, sendo respostas metabólicas para manter a pressão de turgor celular e reduzir a ação de espécies reativas de oxigênio que degradam as membranas celulares (Farooq et al., 2009). Além dessas respostas, os níveis elevados de PAs são considerados marcadores metabólicos em plantas expostas a condições de estresses abióticos, sendo Spd e Spm benéficas para a homeostase de proteínas e desintoxicação de espécies reativas de oxigênio, melhorando a capacidade antioxidante das plantas (Alcázar et al., 2020; Hasan et al., 2021).
As PAs são consideradas moléculas protetoras, além de serem importantes participantes de um complexo sistema de sinalização apresentando uma função vital nos processos de tolerância, dependendo do seu tipo e concentração em condições de estresse (Fazilati & Forghani, 2015; González-Hernández et al. 2022). As PAs estão envolvidas em vários processos metabólicos, incluindo: defesa dos pigmentos fotossintéticos, sistemas antioxidantes, interação hormonal e homeostase iônica, que em última análise contribuem para a adaptação ao estresse, promoção do crescimento e desenvolvimento das plantas (Kaur & Das 2022; Shao et al., 2022).
Portanto, pode-se considerar que os efeitos benéficos do uso de L-aminoácidos nas plantas, em parte, podem ter relação com o metabolismo da PAs, levando em conta que a síntese de seus percussores L-arginina, L-ornitina e L-metionina, assim como de todos os aminoácidos, se conectam em algum ponto com a L-glutamina ou com o L-glutamato (Majumdar et al., 2016) pela ação das aminotransferases, enzimas que atuam para produzir o conjunto dos aminoácidos das plantas de acordo com suas demandas metabólicas (Pratelli & Pilot, 2014).
As PAs são parte de um grupo de metabólitos chamados aminas biogênicas, muito estudadas na qualidade de alimentos e saúde humana, como por exemplo a dopamina, neurotransmissor conhecido como “hormônio da felicidade”, que no metabolismo vegetal tem sido relatada como indutora de tolerância aos estresses abióticos (Ahammed & Li., 2023); e as indolaminas, como a serotonina (5-hidroxytriptamina) também relacionada com a sensação de bem estar; e a triptamina, derivada do aminoácido L-triptofano, ambas compartilhando nas plantas funções com o hormônio auxina (Erland & Saxena., 2019).
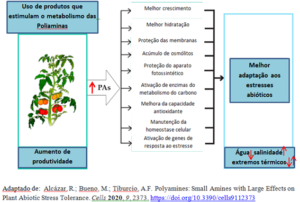
A figura apresenta o resumo gráfico dos potenciais efeitos da aplicação de produtos que estimulam o metabolismo das PAs.